THE EFFECTS OF LEPTIN IN COMBINATION WITH A CANNABINOID RECEPTOR 1 ANTAGONIST, AM 251, OR CANNABIDIOL ON FOOD INTAKE AND BODY WEIGHT IN RATS FED A HIGH-FAT OR A FREE-CHOICE HIGH SUGAR DIET
INTRODUCTION
Food intake and energy balance are controlled by complex neuronal pathways including peripheral and central neurotransmitter systems. Leptin is a key adipokine involved in food intake regulation. This hormone affects multiple neural circuits involved in the control of energy balance (1). Leptin decreases food intake through the inhibition of orexigenic and stimulation of anorectic pathways in the hypothalamic arcuate nucleus and the medullary nucleus of the solitary tract (1-3). Accordingly, leptin has been found to suppress food intake stimulated by ghrelin (4) and to enhance the inhibitory effect of cholecystokinin (CCK) (5), peptide YY (PYY) (6), glucagon-like peptide-1 (GLP-1) and its agonist, exendin-4 (7, 8) on food consumption. Leptin was also demonstrated to contribute to appetite regulation by the inhibition of behavioral responses to palatable, especially sweet and fat foods (9, 10), including sucrose-rich food (11). To date, however, very little has been known about the effect of leptin on the intake of palatable food. Independently of the effect on feeding behavior, the hormone may cause a negative energy balance due to the stimulation of energy expenditure, an event occurring even in leptin-resistant obese animals (12).
The endocannabinoid system is another important regulator of appetite that may be a target for obesity treatment (13). Endocannabinoids stimulate food intake through homeostatic and non-homeostatic mechanisms via the cannabinoid 1 (CB1) receptor (14). Accordingly, CB1 receptor antagonists, such as rimonabant, were shown to reduce appetite and body weight in both obese humans and animals (15). In particular, inactivation of the peripheral CB1 receptor was demonstrated to inhibit the intake of high-carbohydrate and high-fat diet (16) as well as to reduce motivation to work for palatable sucrose pellets (17) or chocolate-flavored food in the rat (18). Similarly, cannabidiol (CBD), a nonpsychoactive cannabinoid found in marihuana, has been recently shown to diminish body weight gain in rats following repeated administration (19), to attenuate hyperphagia induced by the CB1 or 5-HT1A receptor agonists (20) and to contribute to the anti-inflammatory effects of cannabis in the gastrointestinal tract (21). Cannabidiol is known to act either as a CB1 receptor antagonist, CB2 receptor inverse agonist, transient receptor potential vanilloid-1 (TRPV1) agonist, G protein-coupled receptor 55 (GPR55) antagonist or 5-hydroxytryptamine (5HT) 1A and 2A receptor agonist and 5-HT3A receptor antagonist (22).
Interestingly, obesity is characterized by the diminished sensitivity to leptin (23) and overactivity of the endocannabinoid system (24) and there is also a growing body of evidence that endocannabinoids interplay with leptin to affect energy balance. It seems that leptin inhibits tonically the activity of the endocannabinoid system since impaired leptin signaling results in the increased hypothalamic content of endocannabinoids (25), overexpression of CB1 receptor mRNA (26), and enhanced bioelectrical response of hypothalamic neurons involved in food intake regulation to cannabinoids (27) in rodents. A recent report (28) shows that the anorectic response to leptin is mediated entirely by the inhibition of the activity of the hypothalamic endocannabinoid system thus confirming the importance of interactions between leptin and endocannabinoids in food intake control. Leptin was also found to enhance the inhibitory effect of rimonabant on food intake in obese rats (29). However, in that study, rimonabant was administered at a dose that was found to cause drug tolerance on repeated injections. Similarly, leptin dose employed by Boustany-Kari et al. (29), i.e., 500 µg/kg body weight (b.w.), was relatively high compared to that employed in our earlier study (8), where peripherally injected leptin administered at a dose as low as 100 µg/kg was demonstrated to decrease significantly food intake in rats.
Hence, the current study has been aimed at investigations on possible synergistic effects of another CB1 receptor antagonist, AM 251, and leptin administered at low doses on food intake and body weight changes in rats maintained on different high-calorie diets, i.e., a high-fat (HF) and free-choice (FC) high-sucrose diet. Additionally, using the FC paradigm, we have checked whether these drug combinations might affect preferences to highly rewarding sucrose-rich food. Since, to date, there have been no studies on possible interactions between leptin and CBD in terms of their effects on food intake and body weight, such relationship was also tested.
MATERIAL AND METHODS
Animals
All procedures used in this study were reviewed and approved by the Ethics Committee of the Medical University of Lodz.
Male Wistar rats from our breeding stock (Medical University of Lodz, Poland), weighing 250–350 grams were used in the experiments. Each experimental group included 7 to 10 rats. During the experiments all the animals were housed in individual, plastic cages and maintained on a 12:12 hour light-dark cycle (lights off at 6.00 p.m.), at 20–22°C with free access to food and water as described below. Each rat received a preweighed amount (100 g) of food and tap water in a calibrated bottle every day throughout the experiment. The rats were handled every day before the beginning of the experiment to accustom them to experimental conditions.
Dietary composition
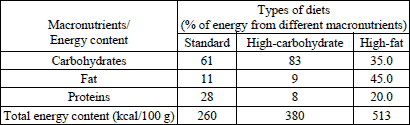
During the experiments, rats were maintained on either 45% HF diet (Ssniff® EF R/M acc. D12451 (I) mod.; ssniff Spezialdiaten GmbH, Soest, Germany) or an FC diet consisting of standard (ST), pelleted rat chow (LSM, AGROPOL, Motycz, Poland) and commercial pellets containing complex carbohydrates, including 35 g sucrose per 100 g. In the free-choice study, both the standard and sweet chow were placed together in a container in the lid of each cage. Since olfactory cues play an important role in food preferences (30), we used chocolate-flavored sweet chow. The nutrient composition and energy content of each diet are shown in Table 1. All types of feed were also enriched with vitamins and minerals.
Chemicals
Leptin (Bachem, Switzerland) was dissolved in 0.9% sterile saline. AM 251 (1-(2,4-dichlorophenyl)-5-(4-methoxyphenyl)-4-methyl-N-(1-piperidinyl)-1H-pyrazole-3 carboxamide; Tocris Bioscience, UK) was initially dissolved in dimethylsulphoxide (DMSO; SIGMA, St. Louis, MO, USA) and then diluted with 0.9% saline using cremophore (TWEEN 80; SIGMA, St. Louis, MO, USA). The final concentrations of DMSO: saline:TWEEN 80 were 2:97:1 (v/v). Cannabidiol (2-[(1R,6R)-3-methyl-6-(1-methylethenyl)-2-cyclohexen-1-yl]-5-pentyl-1,3-benzenediol; Axon Medchem, Groningen, the Netherlands), was dissolved in DMSO and then diluted with saline to the appropriate concentration as indicated above. All drugs, including drug combinations, were administered as a single injection at a volume of 0.1 ml per 100 g b.w.
Leptin was used at a dose of 100 µg/kg which was found previously to suppress standard food consumption (8) and extend the anorectic action of peptide YY (6). AM 251 was injected at a dose of 1 mg/kg that was shown recently to exert a synergistic, inhibitory effect on food intake when administered together with exendin-4 in rats maintained on a FC diet (31). A CBD dose used in our study (3 mg/kg) was comparable to that found previously to reduce body weight in rats (19).
Experimental procedure
The experimental protocol included 4 studies in which the effects of leptin in combination with AM 251 or CBD on the consumption of a HF or FC diet and water as well as body weight changes were tested. At the beginning of the experiments rats were placed in individual cages and switched to a HF or FC diet. They were allowed to accustom to a new diet for 2 days and then the baseline food/water intake were recorded daily at 24 hour intervals for the next 3 days before drug treatment. Thereafter, the rats received drug injections once daily for 3 subsequent days as described below and food/water intake were measured throughout this period 24 hours after each injection. Spilled feed was collected from the bottom of the cages and taken into account when measuring the remaining food every day. In each rat, body weight was measured initially at the beginning of the experiment. Further measurements were made at the end of the 3-day period before drug treatment and, finally, 24 hours after the last drug injection, i.e., 3 days later. The two latter values were then compared with the initial body weight and the differences were referred to as body weight gain. All measurements and injections were performed between 3.30 and 5 p.m.
In the first experiment, HF diet-fed rats, divided into 4 groups, were injected intraperitoneally, once daily with either 0.9% saline, 100 µg/kg leptin, 1 mg/kg AM 251 or 100 µg/kg leptin + 1 mg/kg AM 251 over a 3-day period. Immediately after injections, the animals were placed in their cages with free access to food and water. In the second experiment, rats, maintained on the HF diet as above, were injected with either 0.9% saline, 100 µg/kg leptin, 3 mg/kg CBD or both for 3 days. Two further experiments were performed on rats maintained on a FC diet. In the third and fourth study, the rats were injected with either saline, leptin and AM251 alone and in combination or saline, leptin and CBD alone and in combination as described in experiment 1 and 2, respectively.
Additionally, the basal consumption of normal rat chow was measured in a separate group of rats fed ST diet only. These rats underwent the same experimental protocol, i.e., they were kept in individual cages with free access to ST food and water and the daily food consumption was determined every 24 hours over a 3-day period.
Statistical analysis
Data are presented as the means ± S.E.M. Food and water consumption are expressed as mean daily consumption over the 3-day periods before and after drug treatment. Since rats maintained on an FC diet received two kinds of feed with different energy density, i.e., the ST and high-sucrose diet (see above), food intake in these animals has been expressed both in grams and calories. The results of the experiments were analyzed with statistical software STATISTICA 10 (Statsoft, Cracow, Poland). All the variables (food intake, water intake and body weight) were graphically tested for normal distribution using chi-square test which showed that the data were normally distributed. A one-way analysis of variance (ANOVA) was employed to compare the basal daily food consumption in rats maintained on either the ST, HF or FC diet. The effects of drugs on food and water intake and body weight changes in rats fed high-calorie diets were analyzed using a two-way repeated measures ANOVA. Bonferroni post-hoc multiple comparison test was used to determine differences between groups when significant primary effects (drug × time before and after treatment) were found. The differences between groups were considered to be significant when P≤0.05.
RESULTS
Food intake and body weight changes
The mean basal food consumption in rats assigned to groups maintained on either the ST, HF or FC diet is shown in Fig. 1. A one-way ANOVA revealed that the type of diet affected significantly the basal food intake as expressed in grams and calories (F2,383=4.5, P<0.05 and F2,383=259.76, P<0.001, respectively). Rats maintained on a ST diet and both high-calorie diets consumed similar amounts of food expressed in grams. On the other hand, the caloric intake in animals fed the ST diet was significantly lower than that found in HF or FC diet-fed rats (both P<0.001). Rats presented with the HF diet consumed also significantly more calories than those maintained on the FC diet (P<0.001).
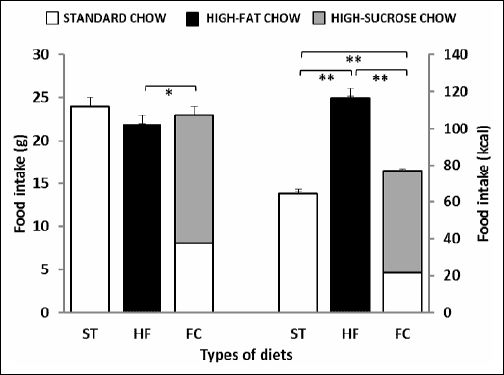 |
Fig. 1. Basal daily intake of standard (ST), high-fat (HF) and free-choice (FC) diet expressed in grams and calories. Data are mean ± S.E.M.; number of animals per group = 6–10. *- P<0.05; **- P<0.001. |
Figs. 2A and 2B present the effects of AM 251 and leptin on food intake and body weight gain, respectively, in rats fed a HF diet. A two-way ANOVA revealed significant main effects of drugs (F3,71=6.3, P<0.001) and time (F1,71=46.71, P<0.001) on the total daily caloric intake in these rats. The post-hoc analysis showed that leptin injected individually decreased markedly (P<0.001) whilst AM 251 did not affect caloric intake compared to baseline values in each group. Co-injection of leptin and AM 251 resulted in a further reduction in food consumption as compared with both leptin alone (P<0.05) and the control group (P<0.01). The two-way ANOVA showed also significant effects of drugs (F3,23=3.59, P<0.05), time (F1,23=4.92, P<0.05) and drugs × time interaction (F3,23=16.87, P<0.001) for body weight. Exposure of the saline-injected controls to the HF diet resulted in the continuous body weight gain (body weight increment after the treatment period was significantly (P<0.001) higher than that measured at the end of the pretreatment period). In contrast, body weight gain in rats treated with leptin or AM 251 injected separately was inhibited because it did not differ significantly from that observed during pre treatment period in both groups of animals. On the other hand, leptin and AM 251 injected together reduced significantly (P<0.01) body weight gain as compared to baseline values in each group. Body weight gain in rats injected with both leptin and the CB1 receptor antagonist was also markedly attenuated in relation to rats treated with leptin only and the controls (both P<0.01).
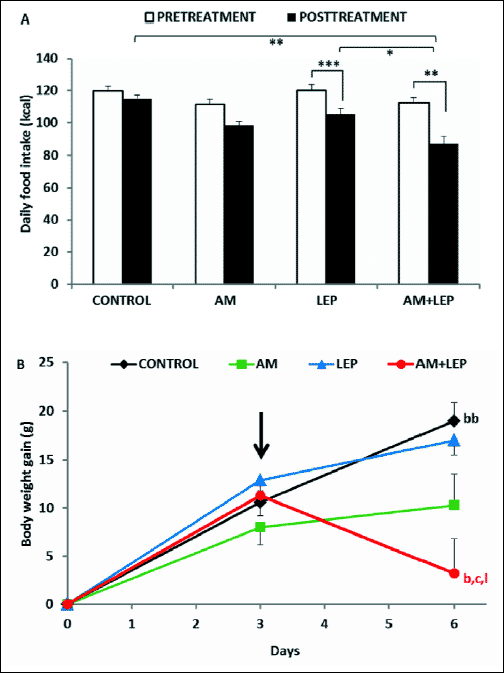 |
Fig. 2. Effects of combined treatment with leptin (LEP) and AM 251 (AM) on the mean daily caloric intake and body weight gain in rats fed a HF diet. Pretreatment (white bars) and post-treatment (black bars) values (panel A) were obtained over the 3-day periods before and after drug treatment, respectively. Body weight (panel B) was measured at the end of the 3-day pretreatment period and then 24 hours after the last injection and compared with the initial body mass obtained on day “0”; the differences were referred to as body weight gain. The arrow indicates the day of the first injection. Significant differences were obtained in relation to basal values in each group (b), saline-injected controls (c) and leptin-injected rats (l). Data are mean ± S.E.M.; number of animals per group = 7–10. *- P<0.05; **- P<0.01; ***- P<0.001; b- P<0.01; bb- P<0.001; c- P<0.01; l- P<0.01. |
Figs. 3A and 3B show the effects of CBD and leptin on food intake and body weight changes, respectively, in rats fed a HF diet. ANOVA performed on these experimental data showed significant main effects of drugs (F3,67=3.54, P<0.05) and time (F1,67=55.77, P<0.001). Leptin and CBD injected either alone or together reduced significantly food consumption when compared to pretreatment values in each group (P<0.001, P<0.01, P<0.01, respectively). There were no significant differences, however, in caloric intake between rats treated concomitantly with leptin and CBD and those treated with each drug injected separately. On the other hand, only rats injected with both leptin and CBD ate significantly (P<0.05) less HF food than the controls. For body weight, ANOVA showed only a significant effect of time (F1,22=95.25, P<0.001). Body weight of the saline-injected control rats (Fig. 3B) increased significantly (P<0.001) during the treatment when compared to pretreatment values. A similar, significant (P<0.01) body weight gain was observed in rats injected with CBD over the same period. On the other hand, body weight increment in leptin-treated rats was comparable with that observed during the pretreatment period and this suppressant effect of leptin on body weight gain was abolished by the simultaneous CBD injection.
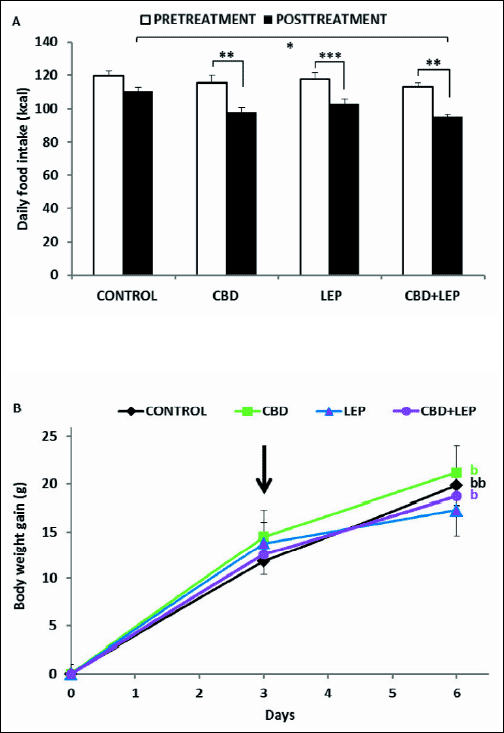 |
Fig. 3. Effects of combined treatment with leptin (LEP) and cannabidiol (CBD) on the mean daily caloric intake and body weight gain in rats fed a HF diet. For explanation see caption to Fig. 2. Significant differences were obtained in relation to basal values in each group (b) only. Data are mean ± S.E.M.; number of animals per group = 7–10. *- P<0.05; **- P<0.01; ***- P<0.001; b- P<0.01; bb- P<0.001. |
Figs. 4 and 5 depict the effects of leptin co-injected with AM 251 or CBD, respectively, on food intake and body weight changes in rats fed an FC diet. In AM 251- and leptin-treated rats there were significant effects of drugs, time and drug x time interaction (F3,110=3.65, P<0.05; F1,110=29.54, P<0.001; F3,110=5.41, P<0.01, respectively) on the total caloric intake. ANOVA showed that there were also significant effects of time (F1,110=26.35, P<0.001) and drug x time interaction (F3,110=3.17, P<0.05) on the total food intake expressed in grams. Rats treated with AM 251 alone or AM 251 together with leptin consumed less food as expressed in grams and there was no significant difference between these two groups (Fig. 4A). Similarly, AM 251 injected alone and together with leptin reduced markedly the total daily caloric intake (both P<0.001) and the difference between these two groups was not significant (Fig. 4B). Regardless whether injected individually or in combination, no drug affected significantly the intake of high-sucrose chow albeit leptin alone tended to increase its consumption. On the other hand, ANOVA revealed that there was a significant main effect of time on ST food consumption expressed both in calories (F1,110=26.05, P<0.001) and grams (F1,110=30.67, P<0.001) in AM 251- and leptin-injected rats on the FC diet. Leptin injected alone or together with AM 251 decreased markedly (both P<0.05) intake of ST chow expressed in calories but not in grams, the difference between these groups was, however, not significant. For body weight, ANOVA revealed a significant effect of drugs x time interaction (F3,29=10.88, P<0.001). Body weight measured at the end of the treatment period tended to increase in each control group of rats maintained on the FC diet as compared to pretreatment values, the differences, however, were not significant (Fig. 4C). AM 251 injected together with leptin reduced significantly body weight gain as compared with both pretreatment values in this group and the control group (both P<0.05).
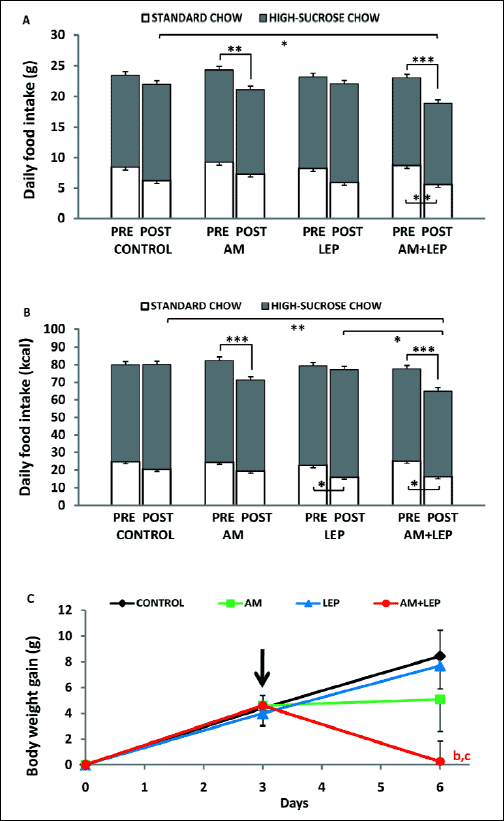 |
Fig. 4. Effects of combined treatment with leptin (LEP) and AM 251 (AM) on the mean daily caloric intake and body weight gain in rats fed a FC diet. Pretreatment (PRE) and post-treatment (POST) values (panels A and B) were obtained over the 3-day periods before and after drug treatment, respectively. Body weight (panel C) was measured at the end of the 3-day pretreatment period and then 24 hours after the last injection and compared with the initial body mass obtained on day “0”; the differences were referred to as body weight gain. The arrow indicates the day of the first injection. Significant differences were obtained in relation to basal values in each group (b) and saline-injected controls (c). Data are mean ± S.E.M.; number of animals per group = 7–10. Data are mean ± S.E.M.; number of animals per group = 7–10. *- P<0.05; **- P<0.01; ***- P<0.001; b- P<0.05; c- P<0.05. |
In rats maintained on an FC diet and injected with leptin and CBD (Fig. 5), ANOVA revealed significant effects of time only for the total daily food intake expressed in calories (F1,113 = 6.22, P<0.05) and grams (F1,113=10.89, P<0.01). For the mean daily consumption of ST food expressed in calories, ANOVA revealed a significant effects of time and drug × time interaction (F1,113 = 24.73, P<0.001; F3,113 = 3.07, P<0.05, respectively). The similar effects were found for the mean daily consumption of ST food expressed in grams (F1,113 = 22.13, P<0.001; F3,113 = 2.21, P<0.05). On the other hand, ANOVA showed no significant effects for high-sucrose food intake. The post-hoc analysis showed that leptin and CBD administered alone or in combination did not affect significantly either the total daily food consumption or high-sucrose chow expressed in calories and grams. CBD and leptin injected separately, however, decreased the consumption of ST chow as compared with baseline intake in these groups (P<0.01 and P<0.05 for values in calories, respectively). This effect was abolished in rats treated concomitantly with both drugs. There were no significant changes in body weight gain in rats injected with leptin and CBD alone or in combination.
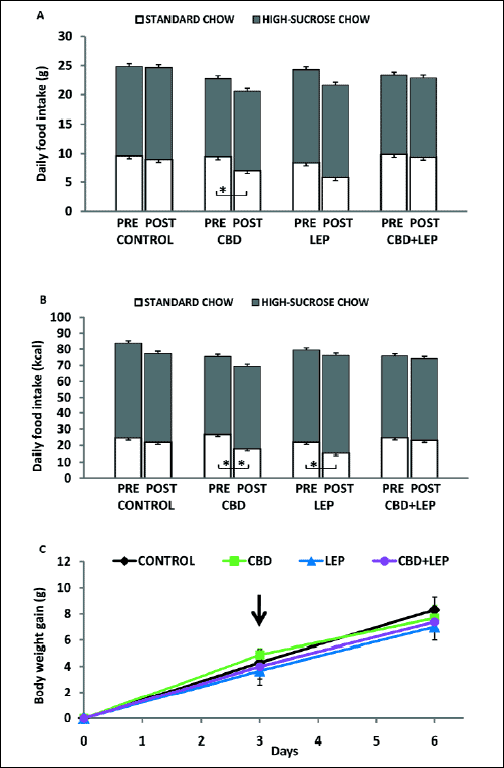 |
Fig. 5. Effects of combined treatment with leptin (LEP) and cannabidiol (CBD) on the mean daily caloric intake and body weight gain in rats fed a FC diet. For explanation see caption to Fig. 4. Data are mean ± S.E.M; number of animals per group = 7–10. Data are mean ± S.E.M.; number of animals per group = 7–10. *- P<0.05; *- P<0.01. |
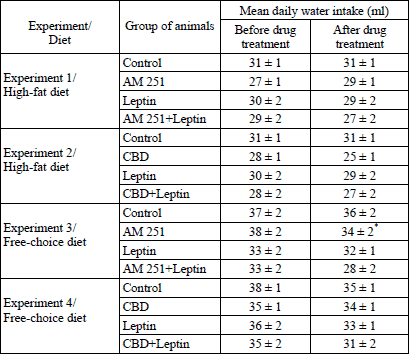
Water intake
The mean daily water intake in all experimental groups is shown in Table 2. A two-way ANOVA revealed significant effects of drugs (F3,112=4.51, P<0.01) and time (F1,112=11.84, P<0.001) on water intake in rats maintained on an FC diet and treated with AM 251 and leptin. In these rats, AM 251 inhibited markedly water consumption as compared to basal values (P<0.01) whilst leptin alone or together with AM 251 had no significant effect on water intake.
DISCUSSION
In the present study, we have extended previous research (29) on interactions between leptin and cannabinoids by examining the effects of combined treatment with leptin and drugs affecting the activity of the cannabinoid system, i.e., a CB1 receptor antagonist, AM 251, and a phytocannabinoid, CBD, on food intake, food preferences, and body weight in rats maintained on a HF or FC diet. Our observation that the basal caloric intake in rats fed a HF or FC diet was significantly greater than that found in rats fed a ST diet only proved that both high-calorie diets were attractive for rats and mimicked conditions leading to overfeeding in humans.
Our finding that leptin injected individually to rats maintained on an FC diet did not affect the total daily calorie intake despite reduced caloric intake from ST chow suggests that there was a “compensatory” increase in sweet chow intake preventing significant changes in the total caloric intake. Indeed, we found that leptin tended to increase the consumption of sweet chow, the differences, however, were not statistically significant. This is in contrast to an earlier report indicating that leptin attenuated behavioral responses to food high in sucrose (17, 32) but is consistent with a recent study demonstrating that the rewarding value of sucrose may be enhanced at low leptin levels (33). On the other hand, we found that leptin did not change either the total daily food, sweet food or standard food intake expressed in grams thus suggesting that the hormone is involved in the control of quality rather than quantity of food consumed. Our finding that leptin did not affect the total daily food intake under FC paradigm is also consistent with results obtained by Harris et al. (34) who found that leptin did not change the total food intake in rats maintained on an FC diet providing that a high-sucrose solution was one of the components of such a diet. On the other hand, they demonstrated that the hormone decreased the total energy intake in rats fed a no-choice, HF diet. Hence, their results matched those obtained in our study. Accordingly to the leptin-dependent effects on energy intake found in our study, the hormone prevented body weight gain in rats on a HF diet but failed to modify body weight in rats on an FC diet.
Our finding that AM 251 injected individually prevented body weight gain in rats on a HF diet is consistent with that reported by Judge et al. (35) at a similar (i.e., 0.83 mg/kg) AM 251 dose. The body weight-reducing effect of AM 251 occurring despite the normal food intake in HF diet-fed rats may be likely attributed to the increased energy expenditure, an event found in rats treated with AM 251 (36). Other authors demonstrated, however, that AM 251 suppressed the intake of both HF and high-carbohydrate diets (35, 37) in the rat. Only the latter finding is consistent with results of the present study. A recent report indicates that AM 251 modulates the endocannabinoid pathways in the rat brain in a diet-specific manner (38) and this might account for differential effects of AM 251 on HF and high-carbohydrate food consumption observed in our study.
In contrast to each drug injected separately, the concomitant injection of leptin and AM 251 resulted in a food intake and body weight reduction both in animals fed a HF and FC diet. Importantly, these effects were achieved using drug doses substantially lower than those employed in a prior study on interactions between leptin and a CB1 receptor antagonist in the control of energy balance in the rat (29). This finding further supports the idea that the combined treatment with anorectic drugs used at low doses may be more beneficial than single drug treatment in terms of effectiveness and, possibly, reduced risk of potential side effects of each drug. The above drug combination did not modify, however, the preference for sweet, high-sucrose diet.
On the other hand, CBD injected individually to rats maintained on a HF diet caused an increase in body weight despite significantly reduced food intake. It seems that this could result from a depressant effect of CBD on the tissue oxygen consumption (39). Likely, CBD decreased energy expenditure and, as a consequence, body weight increased even though energy supply was lower than normal. Previous studies on the effects of CBD on food intake have yielded contradictory results. A number of studies found that CBD administered in a broad dose range up to 80–100 mg/kg had no significant effect on food consumption in mice and rats (40-43) whilst other demonstrated that CBD decreased food intake (44, 45) and attenuated hyperphagia induced by CB1 and 5-HT1A receptor agonists in rats (20). On the other hand, under the FC paradigm, the intake of sucrose solution was found to be much less affected by CBD than the regular chow intake (44), an event occurring also in our study. The data on the effect of CBD on body weight changes are scarce and, again, conflicting. CBD was found to decrease body weight gain in young but increase in aged rats fed a standard but not HF diet (19, 43). Hence, our finding that CBD decreased intake of certain foods but was unable to reduce body weight gain indicates the functional ambiguity of CBD in the modification of energy balance.
In contrast to AM 251, CBD did not modify leptin effects on food consumption, food preferences or body weight changes in FC diet-fed rats and abolished the leptin-induced inhibitory effect on body weight gain in HF diet-fed rats. Hence, no clear benefit of combined use of leptin and CBD on energy balance in rats maintained on a HF or FC diet could be identified in our study. The discrepant effects produced by AM 251 and CBD injected individually or co-injected with leptin on energy balance might result from their ability to modify the activity of different receptors. For example, it is suggested that CBD alters body weight through interaction with the CB2 receptor (19) whilst AM 251-dependent reduction in body weight is attributed mainly to the inhibition of the CB1 receptor (46). Alternatively, differential effects of AM 251 and CBD on energy balance could result from their opposite impact on energy expenditure. AM 251 (36) and leptin (47) were found to increase whilst CBD was shown to decrease (39) oxygen consumption in rats indicating, respectively, a stimulatory or inhibitory effect on energy expenditure. Apparently, simultaneous administration of agents increasing energy expenditure, i.e., leptin and AM 251, resulted in a significant body weight reduction whilst the concomitant administration of agents with the opposite action on metabolic rate, i.e., leptin and CBD, did not affect body weight.
Importantly, the results of our study did not show any significant effects of the tested drug combinations on water intake irrespective of the dietary model. This finding is consistent with our previous results which indicated that AM 251 administered together with exendin-4, a glucagon-like peptide agonist, did not change water consumption although this drug combination reduced both food intake and body weight in rats (48). Moreover, we found that leptin abolished the inhibitory effect of AM 251 on water consumption in rats maintained on an FC diet indicating that the concomitant injection of leptin and AM 251 affects selectively feeding behavior.
In summary, we have shown that the blockade of the CB1 receptor by AM 251 in leptin-treated rats inhibits excessive intake of palatable, high-calorie foods and body weight gain in rats more than would the drugs administered individually. This drug combination does not modify, however, the preference for palatable, high-sucrose chow. In contrast, co-injection of leptin and CBD has not been found to suppress appetite and body weight gain in rats fed high-calorie diets. Since long-term feeding with high-fat high-fructose diet results in cardiovascular and metabolic disorders (49), further investigations should be aimed at examining possible side-effects and effectiveness of the combined treatment with leptin and CB1 receptor antagonists in obese animals fed a high-calorie diet over a longer period of time.
Acknowledgements: This study was supported by the statutory grant 503/6-103-03/503-01 from the Medical University of Lodz.
Conflict of interests: None declared.
REFERENCES
- Jovanovic Z, Yeo GS. Central leptin signalling: beyond the arcuate nucleus. Auton Neurosci 2010; 156: 8-14.
- Sahu A. A hypothalamic role in energy balance with special emphasis on leptin. Endocrinology 2004; 145: 2613-2620.
- Klok MD, Jakobsdottir S, Drent ML. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: a review. Obes Rev 2007; 8: 21-34.
- Ueno N, Dube MG, Inui A, Kalra PS, Kalra SP. Leptin modulates orexigenic effects of ghrelin and attenuates adiponectin and insulin levels and selectively the dark-phase feeding as revealed by central leptin gene therapy. Endocrinology 2004; 145: 4176-4184.
- Barrachina MD, Martinez V, Wang L, Wei JY, Tache Y. Synergistic interaction between leptin and cholecystokinin to reduce short-term food intake in lean mice. Proc Natl Acad Sci USA 1997; 94: 10455-10460.
- Unniappan S, Kieffer TJ. Leptin extends the anorectic effects of chronic PYY(3-36) administration in ad libitumfed rats. Am J Physiol 2008; 295: R51-R58.
- Williams DL, Baskin DG, Schwartz MW. Leptin regulation of the enorexic response to glucagon-like peptide-1 receptor stimulation. Diabetes 2006; 55: 3387-3393.
- Bojanowska E, Nowak A. Interactions between leptin and exendin-4, a glucagon-like peptide-1 agonist, in the regulation of food intake in the rat. J Physiol Pharmacol 2007; 58: 349-360.
- Berridge KC, Ho CY, Richard JM, DiFeliceantonio AG. The tempted brain eats: Pleasure and desire circuits in obesity and eating disorders. Brain Res 2010; 1350: 43-64.
- Fulton S. Appetite and reward. Front Neuroendocrinol 2010; 31: 85-103.
- Domingos AI, Vaynshteyn J, Voss HU, et al. Leptin regulates the reward value of nutrient. Nat Neurosci 2011; 14: 1562-1568.
- Enriori PJ, Sinnayah P, Simonds SE. Garcia Rudaz C, Cowley MA. Leptin action in the dorsomedial hypothalamus increases sympathetic tone to brown adipose tissue in spite of systemic leptin resistance. J Neurosci 2011; 24: 12189-12197.
- Andre A, Gonthier MP. The endocannabinoid system: Its roles in energy balance and potential as a target for obesity treatment. J Biochem Cell Biol 2010; 42: 1788-1801.
- Silvestri C, Di Marzo V. The endocannabinoid system in energy homeostasis and the etiopathology of metabolic disorders. Cell Metab 2013; 17: 475-490.
- Bermudez-Silva FJ, Viveros MP, McPartland JM, Rodriguez de Fonseca F. The endocannabinoid system, eating behavior and energy homeostasis: The end or a new beginning? Pharmacol Biochem Behav 2010; 95: 375-382.
- Randall PA, Vemuri VK, Segovia KN, et al. The novel cannabinoid CB1 antagonist AM 6545 suppresses food intake and food-reinforced behavior. Pharmacol Biochem Behav 2010; 97: 179-184.
- Rasmussen EB, Reilly W, Buckley J, Boomhower SR. Rimonabant reduces the essential value of food in the genetically obese Zucker rat: an exponential demand analysis. Physiol Behav 2012; 105: 734-741.
- Droste M, Saland SK, Schlitter EK, Rodefer JS. AM 251 differentially effects food-maintained responding depending on food palatability. Pharmacol Biochem Behav 2010; 95: 443-448.
- Ignatowska-Jankowska B, Jankowski MM, Swiergiel AH. Cannabidiol decreases body weight gain in rats: involvement of CB2 receptors. Neurosci Letters 2011; 490: 82-84.
- Scopinho AA, Guimaraes FS, Correa FM, Resstel LB. Cannabidiol inhibits the hyperphagia induced by cannabinoid-1 or serotonin-1A receptor agonists. Pharmacol Biochem Behav 2011; 98: 268-272.
- Wallace JL, Flannigan KL, McKnight W, Wang L, Ferraz JG, Tuitt D. Pro-resolution, protective and anti-nociceptive effects of a cannabis extract in the rat gastrointestinal tract. J Physiol Pharmacol 2013; 64: 167-175.
- Campos AC, Moreira FA, Gomes FV, Del Bel EA, Guimaraes FS. Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol in psychiatric disorders. Philos Trans R Soc Lond B Biol Sci 2012; 367: 3364-3378.
- Dardeno TA, Chou SH, Moon HS, Chamberland JP, Fiorenza CG, Mantzoros CS. Leptin in human physiology and therapeutics. Front Neuroendocrinol 2010; 31: 377-393.
- Quarta C, Mazza R, Obici S, Pasquali R, Pagotto U. Energy balance regulation by endocannabinoids at central and peripheral levels. Trends Mol Med 2011; 17: 518-526.
- Di Marzo V, Goparaju SK, Wang L, et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature 2001; 410: 822-825.
- Jelsing J, Larsen P, Vrang N. The effect of leptin receptor deficiency and fasting on cannabinoid receptor 1 mRNA expression in the rat hypothalamus, brainstem and nodose ganglion. Neurosci Lett 2009; 463: 125-129.
- Jo YH, Chen YJ, CChua SC, Talmage DA, Rolel LW. Integration of endocannabinoid and leptin signaling in an appetite-related neural circuit. Neuron 2005; 48: 1055-1066.
- Bermudez-Silva FJ, Cardinal P, Cota D. The role of the endocannabinoid system in the neuroendocrine regulation of energy balance. J Psychopharmacol 2012; 26: 114-124.
- Boustany-Kari CM, Jackson VM, Gibbons CP, Swick AG. Leptin potentiates the anti-obesity effects of rimonabant. Eur J Pharmacol 2011; 658: 270-276.
- Akar F, Mutlu O, Komsuoglu Celikyurt I, et al. Effects of 7-NI and ODQ on memory in the passive avoidance, novel object recognition, and social transmission of food preference tests in mice. Med Sci Monit Basic Res 2014; 20: 27-35.
- Radziszewska E, Wolak M, Bojanowska E. Concurrent pharmacological modification of cannabinoid-1 and glucagon-like peptide-1 receptor activity affects feeding behavior and body weight in rats fed a free-choice, high-carbohydrate diet. Behav Pharmacol 2014; 25: 53-60.
- Figlewicz DP, Bennett JL, Naleid AM, Davis C, Grimm JW. Intraventricular insulin and leptin decrease sucrose self-administration in rats. Physiol Behav 2006; 89: 611-616.
- Domingos AI, Vaynshteyn J, Sordillo A, Friedman JM. The reward value of sucrose in leptin-deficient obese mice. Mol Metab 2014; 3: 73-80.
- Harris RB, Apolzan JW. Changes in glucose tolerance and leptin responsiveness of rats offered a choice of lard, sucrose, and chow. Am J Physiol 2012; 302: R1327-R1339.
- Judge MK, Zhang Y, Scarpace PJ. Responses to the cannabinoid receptor-1 antagonist, AM 251, are more robust with age and with high-fat feeding. J Endocrinol 2009; 203: 281-290.
- Merroun I, Sanchez-Gonzalez C, Martinez R, et al. Novel effects of the cannabinoid inverse agonist AM 251 on parameters related to metabolic syndrome in obese Zucker rats. Metabolism 2013; 62: 1641-1650.
- McLaughlin PJ, Winston K, Swezey L, et al. The cannabinoid CB1 antagonists SR 141716A and AM 251 suppress food intake and food-reinforced behavior in a variety of tasks in rats. Behav Pharmacol 2003; 14: 583-588.
- Rivera P, Luque-Rojas MJ, Pastor A, et al. Diet-dependent modulation of hippocampal expression of endocannabinoid signaling-related proteins in cannabinoid antagonist-treated obese rats. Eur J Neurosci 2013; 37: 105-117.
- Chiu P, Karler R, Craven C, Olsen DM, Turkanis SA. The influence of delta9-tetrahydrocannabinol, cannabinol and cannabidiol on tissue oxygen consumption. Res Commun Chem Pathol Pharmacol 1975; 12: 267-286.
- Fernandes M, Schabarek A, Coper H, Hill R. Modification of delta9-THC-actions by cannabinol and cannabidiol in the rat. Psychopharmacologia 1974; 38: 329-338.
- Wiley JL, Burston JJ, Leggett DC, et al. CB1 cannabinoid receptor-mediated modulation of food intake in mice. Br J Pharmacol 2005; 145: 293-300.
- Riedel G, Fadda P, McKillop-Smith S, Pertwee RG, Platt B, Robinson L. Synthetic and plant-derived cannabinoid receptor antagonists show hypophagic properties in fasted and non-fasted mice. Br J Pharmacol 2009; 156: 1154-1166.
- Ignatowska-Jankowska B, Jankowski MM, Torczynska A. Kurek A, Swiergiel AH. Effects of repeated administration of cannabidiol on body weight gain, food intake and accumulation of adipose tissue in rats. Acta Neurobiol Exp 2011; 71 (Suppl.): 59.
- Sofia RD, Knobloch LC. Comparative effects of various naturally occurring cannabinoids on food, sucrose and water consumption by rats. Pharmacol Biochem Behav 1976; 4: 591-599.
- Farrimond JA, Whalley BJ, Williams CM. Cannabinol and cannabidiol exert opposing effects on rat feeding patterns. Psychopharmacology (Berl) 2012; 223: 117-129.
- Chambers AP, Sharkey KA, Koopman HS. Cannabinoid (CB)1 receptor antagonist, AM 251, causes a sustained reduction of daily food intake in the rats. Physiol Behav 2004; 83: 863-869.
- Wang JL, Chinookoswong N, Yin S, Shi ZQ. Calorigenic actions of leptin are additive to, but not dependent on, those of thyroid hormones. Am J Physiol 2000; 279: E1278-E1285.
- Bojanowska E, Radziszewska E. Combined stimulation of glucagon-like peptide-1 receptor and inhibition of cannabinoid CB1 receptor act synergistically to reduce food intake and body weight in the rat. J Physiol Pharmacol 2011; 62: 395-402.
- Sakr HF. Modulation of metabolic and cardiac dysfunctions by swimming in overweight rats on a high cholesterol and fructose diet: possible role of adiponectin. J Physiol Pharmacol 2013; 64: 231-240.
A c c e p t e d : August 4, 2014